Zmienność przeciwciał
| Ten artykuł od 2007-08 wymaga zweryfikowania podanych informacji. |
Zmienność przeciwciał – występowanie przeciwciał o znacznym stopniu zróżnicowania miejsc wiążących antygen (paratopów). Zmienność przeciwciał jest zjawiskiem nietypowym, bowiem jego podstawą są procesy genetyczne prowadzące do powstania genów całkiem nowych, nieobecnych w linii zarodkowej.
Zmienność przeciwciał wynika z trzech głównych mechanizmów:
- rekombinacji genów – spośród kilkuset genów wybierane są trzy, które łączą się przypadkowo
- tworzenia rejonów P – przed połączeniem genów tworzone są krótkie palindromowe sekwencje bazujące na sekwencjach obecnych w linii zarodkowej
- tworzenia rejonów N – przed połączeniem genów dodawanych jest kilka dodatkowych nukleotydów, tworzących nowe sekwencje, nieobecne w linii zarodkowej.
Oprócz tego, w określonych przypadkach, znaczącą rolę mogą odgrywać mutacje somatyczne oraz konwersja mitotyczna. Pierwszy z tych procesów jest istotny w zjawisku dojrzewania powinowactwa, drugi natomiast bywa głównym źródłem zmienności na przykład u ptaków.
Zmienność kombinacyjna
Najważniejszym zjawiskiem podczas generowania zmienności jest niewątpliwie zmienność kombinacyjna. Z jednej strony stanowi bezpośrednią przyczynę zmienności, z drugiej zaś pozwala na utworzenie rejonów P i rejonów N. Jak więc widać, przedstawiane tutaj mechanizmy nie są od siebie całkowicie niezależne. W rzeczywistości są one jednocześnie przeprowadzane przez jeden kompleks enzymatyczny – rekombinazę – charakteryzujący się istnieniem kilku odrębnych aktywności.
U podstaw zmienności kombinacyjnej leży istnienie wielu różnych genów, które w wyniku łączenia się z sobą tworzą funkcjonalny gen dla części zmiennej przeciwciał. W przypadku łańcucha ciężkiego wyróżnia się kilkadziesiąt genów V, kilkanaście genów D i kilka genów J. Oczywiście niezbędny jest jeszcze DNA kodujący część stałą przeciwciała – jest on zawarty w obrębie genów C. Rozkład genów na chromosomie przedstawia się następująco[a]:
W przypadku zmienności kombinacyjnej dochodzi najpierw do połączenia się genu D z genem J. Zwykle proces ten zachodzi przez wypętlenie fragmentu DNA pomiędzy genem J i genem D użytym do rekombinacji, następnie jest on wycinany. Geny D i J łączą się ze sobą, a wycięty fragment DNA jest zamykany w formę kolistą. Nowe połączenie między genami D i J nosi nazwę złącza kodującego:
J_recombination-diagram2.svg)
W analogiczny sposób powstaje połączenie między nowo utworzonym genem DJ a genem V, przy czym w przypadku, gdy pomiędzy nimi występują inne geny V lub D, są one wycinane i wchodzą w skład kolistego DNA (podobnie rzecz ma się w przypadku tworzenia genu DJ):
J_recombination-diagram3.svg)
W ten sposób powstał gen VDJ, kodujący całą część zmienną przeciwciała. Ostatnim krokiem jest przyłączenie do tego genu fragmentu kodującego część stałą:
J_recombination-diagram4.svg)
Efektem powyższych wydarzeń jest powstanie kompletnego genu dla immunoglobuliny. Może on już podlegać ekspresji, w wyniku której powstają łańcuchy ciężkie przeciwciała. Podobne procesy zachodzą w przypadku łańcucha lekkiego, z tym jednak zastrzeżeniem, że w ich przypadku nie ma genów D - występują tylko geny V, J i C.
Na uwagę zasługuje jeszcze mechanizm, który pozwala na ułożenie genów w odpowiedniej kolejności. Wynika on z pewnych właściwości sekwencji graniczących bezpośrednio z genami i nazywany jest regułą 12/23.
Każdy gen V, D i J posiada z każdej strony układ, na który składają się: heptamer (sekwencja siedmiu nukleotydów) i nonamer (sekwencja dziewięciu nukleotydów), które mają ściśle określony skład nukleotydowy. Pomiędzy nimi natomiast występuje tak zwana wstawka, o długości 12 lub 23 nukleotydów. Reguła 12/23 wynika z tego, że podczas rekombinacji dochodzi do połączenia się tylko tych genów, które mają dwie różne wstawki od strony tworzenia połączenia. Geny V wykazują po stronie 5' wstawkę 12, zaś po stronie 3' wstawkę 23. Geny D po obu stronach mają wstawkę 12, zaś geny J po stronie 5' wykazują obecność wstawki 23, co przedstawione jest poniżej:
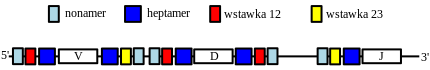
- Podczas połączenia genów dochodzi do wyłączenia nonamerów, heptamerów i wstawek do kolistego DNA (jest to tak zwane złącze sygnałowe).
Gdy dojdzie do rekombinacji obu łańcuchów i ekspresji nowych genów, następuje łączenia się łańcuchów lekkich z ciężkimi (dodatkowo zachodzi jeszcze proces wyłączania allelicznego). W jaki jednak sposób te procesy generują zmienność? Niech przedstawią to poniższe obliczenia:
- W skład genów dla łańcucha ciężkiego przeciwciała wchodzi: 38–46 funkcjonalnych genów V (są także pseudogeny), 23 funkcjonalne geny D i 6 genów J. W związku z tym, w wyniku przypadkowego ich łączenia się, może powstać 42 × 23 × 6 = 5796 wariantów łańcucha ciężkiego.
- Na przykładzie łańcucha lekkiego κ możemy obliczyć, że przy jego 31–35 funkcjonalnych genach V oraz 5 genach J otrzymamy 33 × 5 = 165 wariantów tego łańcucha.
- Łańcuchy ciężkie i lekkie także łączą się ze sobą przypadkowo, dlatego jako końcowy efekt zmienności kombinacyjnej w organizmie może powstać około 5796 × 165 = 956340 wariantów immunoglobulin.
Obliczenia te pokazują także, iż sama zmienność kombinacyjna w przypadku przeciwciał nie daje wystarczającego zabezpieczenia przed infekcjami, gdyż przy znanej obecnie liczbie gatunków pasożytów, bakterii, wirusów i grzybów oraz znacznej często liczbie ich odmian serotypowych, milion wariantów przeciwciał nie jest jeszcze żadną obroną. Ponadto, ze względu na przypadkowość łączenia się genów, wiele swoistości może wiązać patogeny z którymi organizm w ogóle się nie zetknie. Problem ten jest omijany poprzez stworzenie jeszcze większej różnorodności, za pomocą mechanizmów zwanych ogólnie „zmiennością na złączach”.
Zmienność na złączach
Zmienność na złączach jest procesem, w którym dochodzi do zmian sekwencji nukleotydów w obrębie złącza kodującego. Dwa mechanizmy odpowiadające za zmienność na złączach to tworzenie rejonów P oraz N. Do powstania zmienności na złączach dochodzi podczas łączenia się genów. Znaczenie tego rodzaju zmienności jest trudne do przecenienia, zwiększa ona bowiem liczbę wariantów około stukrotnie, czyli z wyjściowego 1 000 000 różnych genów, powstających na skutek zmienności kombinacyjnej, dochodzi do powstania 100 000 000 genów, co daje możliwość odpowiedzi na ogromną liczbę antygenów.
Tworzenie rejonów P
Rejony P, czyli rejony palindromowe, powstają na skutek wykorzystania gotowych genów. Mechanizm jest w tym wypadku następujący:
Rekombinaza przecina DNA, przy czym wypętlony wcześniej fragment cząsteczki jest w tym momencie usuwamy. Wolne końce genów, będące końcami tępymi, zostają zmodyfikowane przez połączenie dwu przeciwbieżnych nici DNA:

Rekombinaza przecina teraz jedną z nici w pewnej odległości od końca, co powoduje powstanie jednoniciowego fragmentu DNA. Do takich miejsc mogą się też dołączać inne nukleotydy (patrz dalej: tworzenie rejonów N), co nie zostało ujęte na rysunku:

Następuje uzupełnienie nowego fragmentu o komplementarne zasady i ligacja DNA:

W ten sposób powstał rejon P. Jak widać, nowo powstałe fragmenty rzeczywiście mają postać palindromów. W powyższym przykładzie: nowo utworzony fragment genu DJ czytany z górnej nici od lewej do prawej ma skład ATGCAT, zaś odczytany wspak tworzy sekwencję czytaną od lewej w nici dolnej.
Tworzenie rejonów N
Rejony N powstają poprzez dodawanie do złącza kodującego dodatkowych nukleotydów. Przypadkowe sekwencje powstają przez działanie transferazy deoksyrybonukleotydów terminalnych. Działanie tego enzymu obaliło jeden z dogmatów genetyki, który zakładał obecność w organizmie jedynie genów pochodzących z linii zarodkowej - w tym wypadku, dzięki dodawaniu, na dodatek przypadkowemu, nukleotydów, powstają zupełnie nowe geny.
Drugim mechanizmem wpływającym na tworzenie nowych genów jest tak zwane chwytanie oligonukleotydów, polegające na dołączaniu krótkich sekwencji, uwalnianych podczas rekombinacji genów, do złącza kodującego.
Inne mechanizmy zmienności przeciwciał
Zmienność może być także uzyskana dzięki mutacjom somatycznym oraz konwersji mitotycznej.
Mutacje somatyczne zachodzą głównie podczas dojrzewania powinowactwa i mają charakter hipermutacji. Konwersja mitotyczna jest natomiast odpowiedzialna za generowanie zmienności tylko u niektórych gatunków zwierząt – u człowieka i innych ssaków jest ona rzadko spotykana, ale u ptaków może stanowić główne źródło zmienności przeciwciał.
Zobacz też
Uwagi
- ↑ Nie pokazano rzeczywistych ilości genów, ale jedynie, w celu uproszczenia, kilka z nich. Jednym kolorem oznaczono kompleksy genów.
Media użyte na tej stronie
Autor:
- V(D)J_recombination-diagram.svg: Wikipedysta:Rantes (png) Marek M (svg)
- derivative work: Marek M (talk)
J_recombination-diagram.svg){kind=link}
Zmienność kombinacyjna
Autor:
- V(D)J_recombination-diagram.svg: Wikipedysta:Rantes (png) Marek M (svg)
- derivative work: Marek M (talk)
Zmienność kombinacyjna

Autor: Wersję rastrową wykonał użytkownik polskiego projektu wikipedii: Rantes, Zwektoryzował: Krzysztof Zajączkowski, Licencja: GFDL
Rysunek do art. zmienność przeciwciał
J_recombination-diagram1.svg)
Autor:
- V(D)J_recombination-diagram.svg: Wikipedysta:Rantes (png) Marek M (svg)
- derivative work: Marek M (talk)
Zmienność kombinacyjna, jednym kolorem oznaczone geny tworzące kompleksy
Autor: Wersję rastrową wykonał użytkownik polskiego projektu wikipedii: Rantes, Zwektoryzował: Krzysztof Zajączkowski, Licencja: GFDL
Rysunek do art. zmienność przeciwciał.
Autor: Wersję rastrową wykonał użytkownik polskiego projektu wikipedii: Rantes, Zwektoryzował: Krzysztof Zajączkowski, Licencja: GFDL
Rysunek do art. zmienność przeciwciał.
Autor:
- V(D)J_recombination-diagram.svg: Wikipedysta:Rantes (png) Marek M (svg)
- derivative work: Marek M (talk)
Zmienność kombinacyjna
Autor: Wersję rastrową wykonał użytkownik polskiego projektu wikipedii: Rantes, Zwektoryzował: Krzysztof Zajączkowski, Licencja: GFDL
Rysunek do art. zmienność przeciwciał.